Research: Leaf Color Function in Plants
The nature and function of leaf colors, particularly in young leaves of tropical plants and senescing leaves in temperate plants.









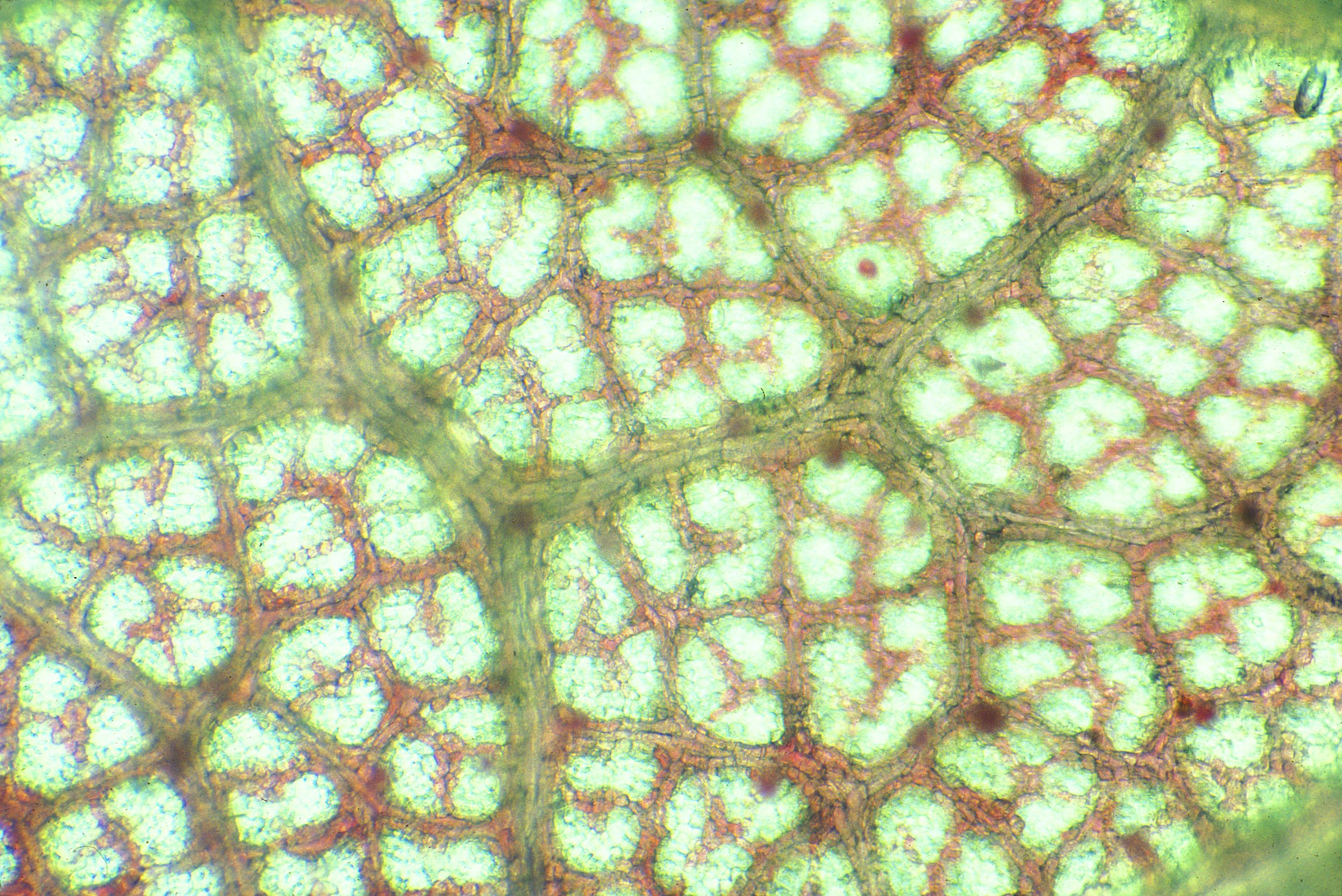



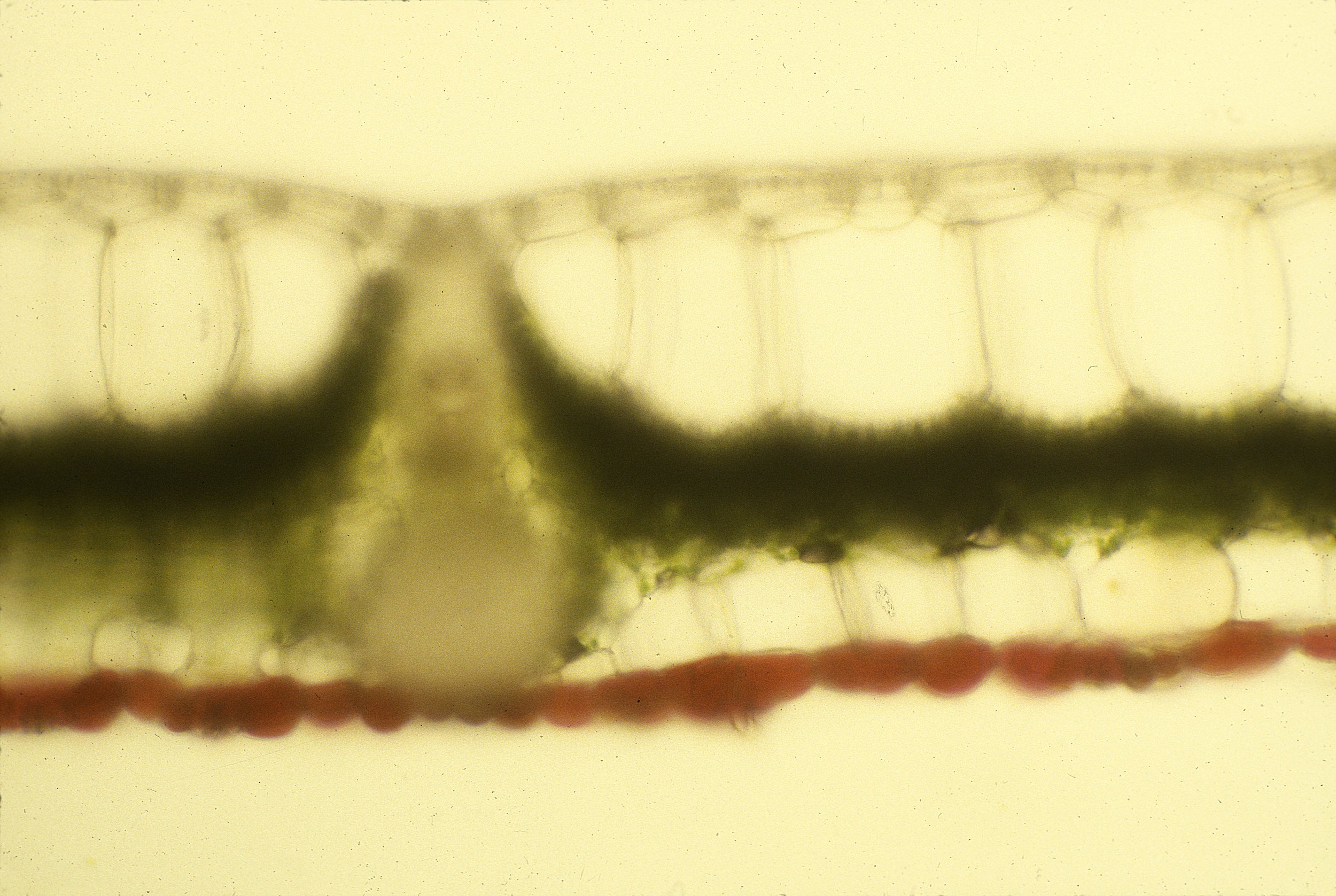





Current Research
-
Present Research on Patterns in Autumn Leaf Color
With good survey data on leaf color (and pigmentation) during leaf senescence from Massachusetts and Colorado, I contacted potential collaborators and came up with a team and a dramatically increased survey, both in totals of species and locations. The collaborators are seen in the document below. With the survey complete, and the write-up under the leadership of Niky Hughes and Mason Heberling, we have written up an abstract for a talk or poster at Botany 2022, meeting this July in Anchorage. After that, we’ll prepare a manuscript and submit it for publication.
Important Articles
-
Unraveling the evolution of autumn colours: an interdisciplinary approach
by Archetti, M., T. F. Döring, S.B. Hagen, N.M. Hughes, S.R. Leather, D.W. Lee, S. Lev-Yadun, Y. Manetas, H.J. Ougham, P.J. Schaberg, and H. Thomas.
in Trends in Ecology and Evolution 24.3, 2009
Leaf colour change is commonly observed in temperate deciduous forests in autumn. This is not simply a side effect of leaf senescence, and, in the past decade, several hypotheses have emerged to explain the evolution of autumn colours. Yet a lack of crosstalk between plant physiologists and evolutionary ecologists has resulted in slow progress, and so the adaptive value of this colour change remains a mystery. Here we provide an interdisciplinary summary of the current body of knowledge on autumn colours, and discuss unresolved issues and future avenues of research that might help reveal the evolutionary meaning of this spectacle of nature.
-
Pigment dynamics and autumn leaf senescence in a New England deciduous forest
by David W. Lee, J. O’Keefe, N.M. Holbrook and T.S. Field
in Ecological Research 18, 2003
The leaves of woody plants at Harvard Forest in Central Massachusetts, USA, changed color during senescence; 70% (62/89) of the woody species examined anatomically contained anthocyanins during senescence. Anthocyanins were not present in summer green leaves, and appeared primarily in the vacuoles of palisade parenchyma cells. Yellow coloration was a result of the unmasking of xanthophyll pigments in senescing chloroplasts. In nine red-senescing species, anthocyanins were not detectable in mature leaves, and were synthesized de novo in senescence, with less than 20 µg cm² of chlorophyll remaining. Xanthophyll concentrations declined in relation to chlorophyll to the same extent in both yellow- and red-leaved taxa. Declines in the maximum photosystem II quantum yield of leaves collected prior to dawn were only slightly less in the red-senescing species, indicating no long-term protective activity. Red-leaved species had significantly greater mass/area and lower chlorophyll a/b ratios during senescence. Nitrogen tissue concentrations in mature and senescent leaves negatively correlated to anthocyanin concentrations in senescent leaves, weak evidence for more efficient nitrogen resorption in anthocyanic species. Shading retarted both chlorophyll loss and anthocyanin production in Cormus alternifolia, Acer rubrum, Acer saccharum, Quercur rubra and Viburnum alnifolium. It promoted chlorophyll loss in yellow-senescing Fagus grandifolia. A reduced red : far-red ratio did not affect this process. Anthocyanins did not increase leaf temperatures in Q. rubra and Vaccinium corymbosum on cold and sunny days. The timing of leaf-fall was remarkably constant from year to year, and the order of senescence of individual species was consistent.
-
Anthocyanins in leaves: distribution, phylogeny and development
by David W. Lee
in Advances in Botanical Research 37, 2002
ABSTRACT
Red pigments, product of different metabolic pathways, occur in terrestrial plants. The flavonoid pathway contributes the greatest diversity, culminating in the prevalence of anthocyanins in the angiosperms. Anthocyanins are produced in flowers and fruits, and also in vegetative organs, but have been poorly researched in the latter. Anthocyanins are commonly produced in:
rapidly expanding leaves of tropical plants;
senescing leaves of temperate plants;
undersurfaces of floating leaves of aquatic plants;
abaxial surfaces of leaves of understory plants; and
leaves subjected to various environmental stresses.
The distribution of anthocyanins in leaves, both in presence and in tissue distribution, is influenced by both phylogeny and development. Few species product anthocyanins in leaf tissues derived from both dermal and ground embryonic tissue. These influences will be important in resolving the ecological roles of anthocyanins in leaves.
-
Anthocyanins in leaves and other vegetative organs: an Introduction
by David W. Lee and Kevin S. Gould
in Advances in Biological Research 37, 2002
ABSTRACT
Although anthocyanins are most recognized as pigments contributing to coloration in fruits and flowers, they are also present in leaves and other vegetative organs. Although their presence has long been recognized, particularly because of their contribution to autumn coloration, the phenomenon has been poorly studied and is not well understood. In this chapter we review the history of research on anthocyanins in leaves, emphasizing the flurry of research at the end of the 19th century as well as the growing body of contemporary research on the topic. We emphasize the various hypotheses of anthocyanin function that were mainly developed more than a century ago, and emphasize recent research that takes advantage of our dramatically increased understanding of whole plant physiology.
-
Anthocyanins in autumn leaf senescence
by David W. Lee
in Advances in Botanical Research 37, 2002
Anthocyanins are synthesized during leaf senescence in certain plants across virtually all biomes, but are most spectacular in the autumn foliage of temperate deciduous forests. The patterns of color production in senescing foliage depend at least partly upon species composition and their phenology. Both ecological and physiological explanations have been raised to explain why plants produce this pigment just before leaf fall. Physiological explanations, as photoprotection, predict that cyanic leaves would be better able to resorb nitrogen during the process of chlorophyll degradation. Ecological explanations predict better dispersal of propagules advertised by association with the brilliantly colored leaves (plausible for only a minority of species), or warning against egg-laying activity of herbivorous insects, as aphids. These hypotheses make predictions that we now can test, to help us understand this old mystery — and majestic phenomenon.
-
Why Leaves Turn Red in Autumn. The role of anthocyanins in senescing leaves of red-osier dogwood
by Taylor S. Feild, David W. Lee, and N. Michele Holbrook
in Plant Physiology 127, 2001
Why the leaves of many woody species accumulate anthocyanins prior to being shed has long puzzled biologists because it is unclear what effects anthocyanins may have on leaf function. Here, we provide evidence for red-osier dogwood (Cornus stolonifera) that anthocyanins form a pigment layer in the palisade mesophyll layer that decreases light capture by chloroplasts. Measurements of leaf absorbance demonstrated that red-senescing leaves absorbed more light of blue-green to orange wavelengths (495-644 nm) compared with yellow-senescing leaves. Using chlorophyll a fluorescence measurements, we observed that maximum photosystem II (PSII) photon yield of red-senescing leaves recovered from a high-light stress treatment, whereas yellow senescing leaves failed to recover after 6 h of dark adaptation, which suggests photo-oxidative damage. Because no differences were observed in light response curves of effective PSII photon yield for red- and yellow-senescing leaves, differences between red- and yellow-senescing cannot be explained by differences in the capacities for photochemical and non-photochemical light energy dissipation. a role of anthocyanins as screening pigments was explored further by measuring the responses PSII photon yield to blue light, which is preferentially absorbed by anthocyanins, versus red light, which is poorly absorbed. We found that dark-adapted PSII photon yield of red-senescing leaves recovered rapidly following illumination with blue light. However, red light induced a similar, prolonged decrease in PSII photon yield in both red- and yellow-senescing leaves. We suggest that optical masking of chlorophyll by anthocyanins reduces risk of photo-oxidative damage to leaf cells as they senesce, which otherwise may lower the efficiency of nutrient retrieval from senescing autumn leaves.
-
Phylogenetic and ontogenetic influences on the distribution of anthocyanins and betacyanins in leaves of tropical plants
by David W. Lee and Timothy M. Collins
in International Journal of Plant Science 162, 2001
We examined the anatomy of expanding, mature and senescing leaves of tropical plants for the presence of red pigments: anthocyanins and betacyanins. We studied 463 species in total, 370 genera, belonging to 94 families. This included 21 species from five families in the Caryophyllales, where betacyanins are the basis for red color. We also included 14 species of ferns and gymnosperms in seven families and 29 species with undersurface coloration at maturity. We analyzed 399 angiosperm species (74 families) for factors (especially developmental and evolutionary) influencing anthocyanin production during expansion and senescence. During expansion, 44.9% produced anthocyanins and only 13.5% during senescence. AT both stages, relatively few patterns of tissue distributions developed, primarily in the mesophyll, and very few taxa produced anthocyanins in dermal and ground tissue simultaneously. Of the 35 species producing anthocyanins both in development and senescence, most had similar cellular distributions. Anthocyanin distributions were identical in different developing leaves of three heteroblastic taxa. Phylogeny has influenced the distribution of anthocyanins in the epidermis and mesophyll of expanding leaves and the palisade parenchyma during senescence, although these influences are not strong. Betacyanins appear to have similar distributions in leaves of taxa within the Caryophyllales and, perhaps, similar functions. The presence of anthocyanins in the mesophyll of so many species is inconsistent with the hypothesis of protection against UV damage or fungal pathogens, and the differing tissue distributions indicate that the pigments may function in different ways, as in photoprotection and free-radical scavenging.
-
Why leaves are sometimes red
by David W. Lee
in Nature 378, 1995
Anthocyanins are present in leaves of autumn foliage and rapidly developing shoots in tropical trees, but their function has never been satisfactorily explained. In addition, leaf undersurfaces of many tropical rainforest understory herbs are also red because of the presence of anthocyanins. We have examined leaves of two such taxa and report that anthocyanins intercept quanta otherwise absorbed by chlorophyll b, thereby protecting leaves from photoinhibition. We thus confirm what may be a general explanation for anthocyanin function in leaves.
-
Young Leaf Anthocyanin and Solar Ultraviolet
by David W. Lee and J. B. Lowry
in Biotropica 12.1, 1980
ABSTRACT
For six tropical rainforest species, young leaves produced on rapidly flushing shoots had markedly lower reflectance in the UV-B region than did mature leaves and contained higher levels of anthocyanins and total phenols. Progressive changes in these characteristics provide empirical support for an earlier suggestions that anthocyanins in flushing shoots of tropical trees have adaptive value through their ultraviolet absorption.
-
The selective advantages of anthocyanins in developing leaves of mango and cacao
by David W. Lee, Susan Brammeier, and Alan P. Smith
in Biotropica 19.1, 1987
ABSTRACT
Although the developing leaves of mango and cacao contain appreciable concentrations of anthocyanins not present in mature leaves, these compounds are a small proportion of the total phenolic concentration. For this and other reasons anthocyanins do not seem to be important in the developing leaves as: (1) a screen against ultraviolet radiation; (2) a mechanism for elevating leaf temperature: (3) a means of defense or aposematic coloration against herbivory; and (4) a part of any postulated physiological mechanism, as photosynthesis. Anthocyanic coloration may be a by-product of the metabolism of other flavonoid compounds in these rapidly growing organs.
-
Abaxial anthocyanin layer in leaves of tropical rain forest plants: enhancer of light capture in deep shade
by David W. Lee, J.B. Lowry, and B. C. Stone
in Biotropica 11.1, 1979
The permanent pigmentation of the leaves of tropical rain forest herbs with anthocyanin has traditionally been viewed as a mechanism for enhancing transpiration by increased heat absorption. We report measurements to ±0.1°C on four Indo-malesian forest species polymorphic with respect to color. There were no detectable differences in temperature between cyanic and green leaves. In deeply shaded habitats, and temperature difference would arise from black-body infrared radiation which all leaves absorb and to which anthocyanins are transparent. Reflectance spectra of the lower leaf surfaces of these species revealed increased reflectance around 650-750 nm for cyanic leaves compared with green leaves of the same species. In all species anthocyanin was located in a single layer of cells immediately below the photosynthetic tissue. These observations provide empirical evidence that the cyanic layer can improve photosynthetic energy capture by back-scattering additional light through the photosynthetic tissue.